{kind=link}
{kind=link}
{kind=link}
{kind=link}
CMIP5模式对未来升温情景下全球陆地生态系统净初级生产力变化的预估
[朱再春 , 刘永稳, 刘祯, 朴世龙]
, 刘永稳, 刘祯, 朴世龙]
, 刘永稳, 刘祯, 朴世龙]
|
|


作者简介:朱再春,男,博士,zhu.zaichun@pku.edu.cn
针对《巴黎协定》提出的温控目标,利用耦合模式比较计划第五阶段(CMIP5)模式在RCP2.6、RCP4.5和RCP8.5情景下的模拟结果,初步分析了全球升温情景下陆地生态系统净初级生产力(NPP)相对于参考时段(1986—2005年)的变化,重点分析了1.5℃和2℃升温时NPP相对于参考时段的变化量,并探讨了大气CO2浓度、气温、降水和辐射的变化及其对NPP变化的影响。CMIP5基于各典型浓度路径模拟的全球陆地生态系统NPP均呈增加趋势,且NPP增加量与升温幅度成正比。在相同的升温幅度下,基于各典型浓度路径模拟的各环境因子和NPP的变化量较为一致。陆地生态系统NPP总量增加主要由大气CO2浓度上升驱动,其他环境因子的影响相对较弱。中国东南部、非洲中部、美国东南部和亚马孙雨林西部地区NPP增加最明显。NPP变化量的空间格局主要由大气CO2浓度增加和升温控制,降水和辐射的影响相对较小。具体而言,大气CO2浓度上升对中低纬度的NPP变化贡献最大,对北方高纬度地区NPP变化贡献较小。温度上升有利于促进北方高纬度地区和青藏高原地区NPP,但对中低纬度地区的NPP有较强的抑制作用。鉴于既有典型浓度路径和地球系统模型的限制,本文对未来升温情景下陆地生态系统NPP的预估仍存在较大的不确定性,需要在未来的研究中进一步改进。
We tentatively analyzed differences between global terrestrial net primary productivity (NPP) under global warming by the targeting temperature of the Pairs Agreement and that during 1986-2005. We addressed the changes in global terrestrial NPP, changes inrelating environmental factors (atmospheric CO2 concentration, temperature, precipitation, and radiation), and their contribution to the NPP changes under global warming of 1.5℃ and 2℃. The projected global terrestrial NPP increases in proportion to the warming magnitude based on the results from CMIP5 models that runs under the three representative concentration pathways (RCP2.6, RCP4.5 and RCP8.5). Changes in the projected environmental factors and NPP at given warming magnitude are generally consistent across the three RCPs. The increasing atmospheric CO2 concentration is the dominant factor that drives the total amount of global terrestrial NPP, while the contributions of other environmental factors are relatively small. The most notable increases in NPP locate in southeast China, central Africa, southeast U.S. and western Amazonia. The spatial pattern of NPP changes are mainly controlled by atmospheric CO2 concentration increase and warming, while precipitation and radiation contribute much fewer. The effects of increasing atmospheric CO2 concentration on NPP are stronger at lower latitudes but weaker at northern high latitudes. Warming benefits ecosystem NPP at northern high latitudes and Tibetan Plateau but strongly depletes ecosystem NPP at lower latitudes. Our analyses of global terrestrial ecosystem NPP changes under future global warming scenarios still have significant uncertainties due to limitations of current RCPs and earth system models, which needs further refinements.
引 言
陆地生态系统通过光合作用等吸收大气中的CO2, 是重要的碳汇[1, 2, 3, 4, 5]。研究表明, 陆地生态系统能够吸收相当于人类活动排放量1/3的CO2, 对降低大气CO2浓度的上升速率和缓解气候变暖具有重要作用[3, 6]。在陆地生态系统碳循环过程中, 净初级生产力(NPP)是指单位时间和单位面积上绿色植物通过光合作用所积累的有机干物质总量, 是总初级生产力中减去自养呼吸消耗之后的剩余部分, 是包括人类在内的几乎所有生命有机体的物质和能量的基础[3, 7, 8]。NPP是生态系统碳循环的起点, 是海洋-陆地-大气物质交换最重要的通量之一[9], 其变化会直接影响大气CO2浓度, 从而对气候变化产生重要影响[6, 10]。同时, NPP对气候变化十分敏感, 地球系统未来气候变化对全球NPP总量和空间分布具有重要作用[11]。因此, 研究未来气候条件下陆地生态系统NPP的总量及空间格局变化, 对预测和评估未来全球碳循环对人类社会发展的影响以及制定科学的气候政策具有重要意义。
《巴黎协定》正式确定了较之于工业化前, 21世纪末全球平均气温升高控制在2℃以内的目标, 并首次将1.5℃温控目标确立为应对气候变化的长期努力方向, 为全球绿色发展和低碳转型指明了方向, 是全球气候治理的里程碑[12, 13]。IPCC AR5基于大量低浓度排放情景对2℃目标的可行性和实现路径进行了详细论证, 但对1.5℃目标的论述十分有限。对此, 联合国气候变化框架公约委托IPCC于2018年发布关于全球升温1.5℃的特别报告[14]。这将促进学界就全球升温1.5℃对自然生态系统和人类社会经济的影响进行系统评估[13, 14, 15, 16]。然而, 《巴黎协定》仅阐述了未来全球平均气温升高的控制目标与途径, 而目前却无明确的排放情景和相应的模型模拟结果。因此, 本文拟以耦合模式比较计划第五阶段(CMIP5)的地球系统模型在典型浓度路径(RCPs)下的模拟结果, 对全球升温情景下陆地生态系统NPP的变化进行前瞻性的分析。具体而言, 本研究首先分析RCP2.6、RCP4.5、RCP8.5情景下的全球升温对陆地生态系统NPP的影响, 着重比较全球升温1.5℃和2℃时陆地生态系统NPP相对于参考年份(1986— 2005年)的不同变化, 再通过分析大气CO2浓度、气温、降水、辐射等环境因子的变化对NPP变化的影响, 初步探讨在《巴黎协定》提出的温控目标下陆地生态系统NPP对未来气候变化的响应机制。
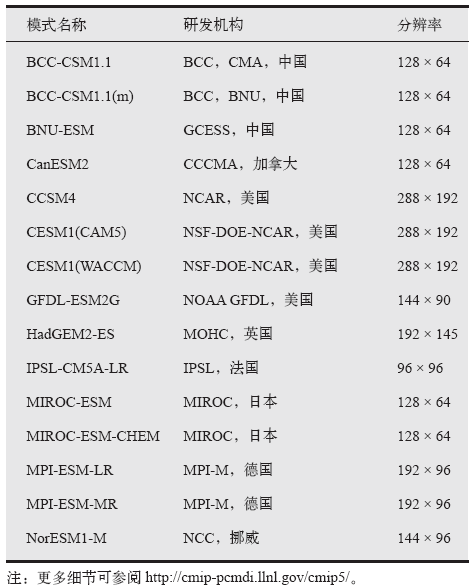
本文首先筛选出CMIP5中同时提供Historical、RCP2.6、RCP4.5和RCP8.5情景下模拟的1861— 2099年逐月NPP、温度、降水、辐射数据的所有地球系统模式(表1), 并将其模拟的逐月NPP、温度、降水和辐射数据分别处理为年总NPP、年均温度、年总降水和年均辐射。由于地球系统模式输出变量的空间分辨率不同, 本文采用一阶守恒重采样方法将所有模型数据统一重采样为1° × 1° [17]。
| 表1 15个CMIP5地球系统模式的基本特征 Table 1 The characteristics of 15 CMIP5 models used in this study |
基于15个地球系统模式的全球年均温模拟数据, 本研究选取1861— 1890年全球平均年均温度作为全球升温的参考温度, 从1890年开始向后搜索首次满足9年平均年均温超过参考温度一定升温幅度(如1.5℃)的年份区间, 并定义该年份区间的中间年为达到该升温幅度的年份。分别计算1986— 2005年平均年总NPP、年均大气CO2浓度、年均温度、年总降水量和年均辐射量作为当前NPP和相应的环境变量的参考值。首次达到某升温幅度的9年年份区间内的平均年总NPP和平均环境变量与当前参考值之间的差值定义为达到该升温幅度时NPP和环境因子相对于当前状态的年变化量。
为确定各环境因子对NPP变化的贡献, 本文首先利用公式(1)计算NPP对各环境因子的敏感性(N)。其中β 、a、b、c分别为NPP对大气CO2浓度(C)、温度(T)、降水(P)和辐射(R)的敏感性, d和分别为回归常数项和残差。
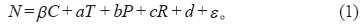
然后计算各环境因子从参考年份至全球升温超过某升温幅度的年份区间的年变化量, 各环境因子的变化量与敏感性的乘积即为各环境因子对NPP变化的贡献。
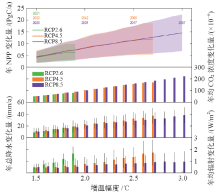
CMIP5模式模拟结果表明, 全球陆地生态系统NPP总量随着全球升温幅度的增加而增加(图1)。全球升温1.5℃时, RCP2.6(2021年)、RCP4.5(2022年)和RCP8.5(2020年)情景对应的陆地生态系统NPP相对于参考时段(1986— 2005年)的年增加量分别为4.17± 2.47 PgC/a、4.55± 2.57 PgC/a和4.24± 2.43 PgC/a(均值± 方差)。仅在RCP4.5(2042年)和RCP8.5(2034年)情景下, 全球平均温度能够升高2℃。两种情景下全球NPP的年增量分别为8.32± 4.68 PgC/a和8.04± 4.52 PgC/a。仅在RCP8.5(2057年)情景下, 全球升温幅度能达到3℃, 其所对应的NPP年增量为14.60± 7.76 PgC/a。
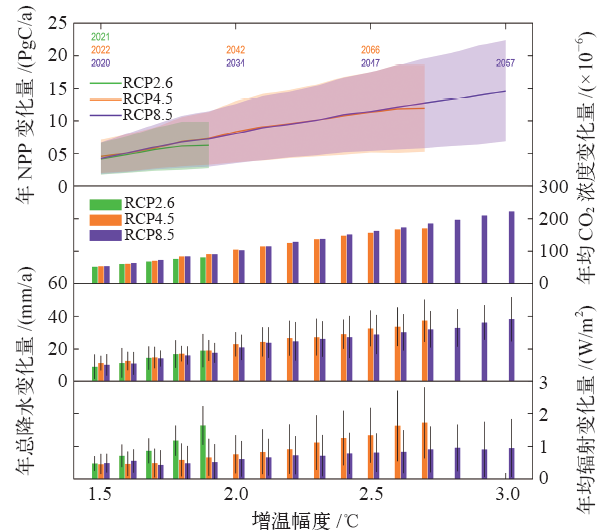 | 图1 全球陆地生态系统NPP、大气CO2浓度、降水和辐射随全球升温的变化注: 图中, 绿色、橙色和紫色数字分别代表RCP2.6、RCP4.5 和RCP8.5 情景下达到一定增温幅度(1.5℃、2℃、2.5℃和3℃)的年份。绿色、橙色和紫色阴影以及黑色误差条分别代表RCP2.6、RCP4.5 和RCP8.5 情景下CMIP5 各模式模拟结果的不确定性范围(标准差)。Fig. 1 Changes in global terrestrial ecosystem NPP, atmospheric CO2concentration, precipitation and radiation with future global warming |
除温度外, 大气CO2浓度、降水、辐射也是影响陆地生态系统NPP变化的主要环境因子[2]。分析这些环境因子未来的变化有助于揭示陆地生态系统NPP未来变化的驱动机制。基于CMIP5多模式平均结果表明, 在不同的升温幅度下, 大气CO2浓度的增量与全球升温幅度成正比, 这与“ 温室效应” 理论一致[3]。全球升温1.5℃时, 模式在RCP2.6、RCP4.5和RCP8.5情景下的模拟结果均表明全球地表植被地区年降水量变化不显著, 但各模式间差异较大(RCP2.6:8.90± 7.62 mm/a, RCP4.5:11.28± 4.45 mm/a, RCP8.5:10.08± 6.73 mm/a)。随着升温幅度的增加, 全球陆地生态系统的降水量也在不断增加。全球升温2℃时, 模式模拟的全球陆地生态系统平均年总降水量较之于参考时段分别增加22.76± 7.40 mm/a(RCP4.5)、20.84± 7.82 mm/a(RCP8.5), 全球升温3℃时, 平均年总降水量增加38.14± 13.58 mm/a(RCP8.5)。与大气CO2浓度、降水相似, 辐射增量在各典型浓度路径下的模式平均值也随着增温幅度的增加而递增, 但模式间差异较大。升温1.5℃时, 年均辐射增量分别为0.48± 0.23 W/m2(RCP2.6)、0.46± 0.31 W/m2(RCP4.5)、0.49± 0.28 W/m2(RCP8.5); 升温2℃时, 年均辐射增量分别为0.76± 0.59 W/m2(RCP4.5)、0.62± 0.55 W/m2(RCP8.5); 升温3℃时, 年均辐射增量为0.95± 0.89 W/m2(RCP8.5)。
大气CO2、温度、降水、辐射等环境因子通过复杂的生物物理过程驱动全球陆地生态系统NPP的变化[2, 18, 19]。本文通过研究全球陆地生态系统NPP对各环境因子的敏感性和各因子的变化量, 进一步揭示各环境因子在全球升温情景下全球陆地生态系统NPP增加中的贡献。自1986年至全球升温1.5℃的年份, CMIP5在3种典型浓度路径下的模拟结果表明大气CO2浓度、温度、降水和辐射对全球陆地生态系统NPP增加量(4.17~4.55 PgC/a)的贡献分别为4.61~4.89 PgC/a、-0.91~-0.46 PgC/a、0.47~0.48 PgC/a和-0.15~-0.08 PgC/a。自1986年至全球升温2℃的年份, 在RCP4.5和RCP8.5情景下, 大气CO2浓度、温度、降水和辐射对全球陆地生态系统年总NPP增加量(8.04~8.32 PgC/a)的贡献分别为6.82 PgC/a、0.82 PgC/a、1.15 PgC/a和-0.35 PgC/a(RCP4.5); 7.23 PgC/a、0.20 PgC/a、0.94 PgC/a和-0.20 PgC/a(RCP8.5)。
2.2 1.5℃和2℃升温情景下NPP及各环境因子变化的空间格局
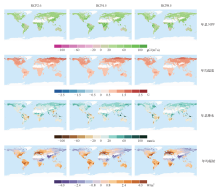
陆地生态系统NPP在全球升温1.5℃的情景下相对于参考时段NPP的变化量的空间格局具有明显的异质性(图2)。在3种典型浓度路径下的模拟的陆地生态系统NPP变化量的空间格局比较一致, 超过95%的区域NPP呈增加趋势。NPP增量较大(> 40 gC/(m2∙ a)的区域主要分布于中国东南部、非洲中部、美国东南部和亚马孙雨林西部地区, 而NPP减少的区域零星分布于北美洲中部、亚马孙雨林东北部、非洲南部和澳大利亚地区。
 | 图2 全球升温1.5℃时CMIP5模式在RCP2.6、RCP4.5和RCP8.5情景下预估的全球年总NPP、年均温度、年总降水、年均辐射相对于参考时段(1986— 2005年)的变化量的空间分布注:白色区域是非植被区域, 主要是沙漠和冰雪覆盖地区。Fig. 2 Spatial pattern of changes in annual total NPP, annual mean temperature, annual total precipitation and annual mean down welling shortwave radiation under global warming of 1.5℃ relative to the reference period 1986 to 2005 based on CMIP5 simulations under RCP2.6, RCP4.5 and RCP8.5 |
全球平均升温1.5℃情况下, 北方高纬度地区升温幅度最明显(> 1.0℃), 中低纬度升温幅度相对较小(0.5~1.0℃)(图2)。全球升温格局在3种典型浓度路径下差异并不明显。降水减少最多的区域主要分布于亚马孙雨林、非洲南部和澳大利亚地区, 增加最多的区域则主要分布于北方高纬度地区、非洲中部和东南亚地区。短波下行辐射增加最明显的区域为美国东部、亚马孙地区、欧洲和非洲南部, 而辐射降低的区域则分布于俄罗斯东北部、印度和东非地区。
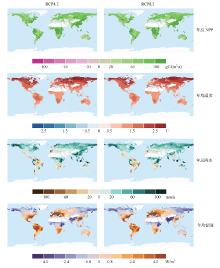
CMIP5模式平均模拟结果仅在RCP4.5和RCP8.5情景下升温能够达到2℃(图3)。全球升温2℃时, 陆地生态系统NPP几乎在所有区域均相对于参考时段状态有所增加, NPP降低的面积不超过全球植被面积的2%。年总NPP增加量最多(> 100 gC/(m2∙ a))的区域分布于中国南部、非洲中部、美国东部和亚马孙西部地区。与升温1.5℃相比, 全球升温2℃时气候因子的变化更加明显。年均温度变化较为明显的区域依然是北方高纬地区(> 2.0℃)。降水增加的区域主要分布于北方高纬度地区以及非洲中部和东部地区。升温2℃时, RCP4.5情景下的模拟结果表明中国南部地区年总降水变化量将会明显增多(60 mm/a), RCP8.5情景下模拟的该地区的降水量则增加相对较小甚至稍有降低。辐射量明显增加的区域主要分布于中国东南部、欧洲、美国东部沿海地区、亚马孙地区和非洲南部, 而辐射量降低的区域则主要分布于印度、北方高纬度地区以及非洲中部和东部的部分地区。
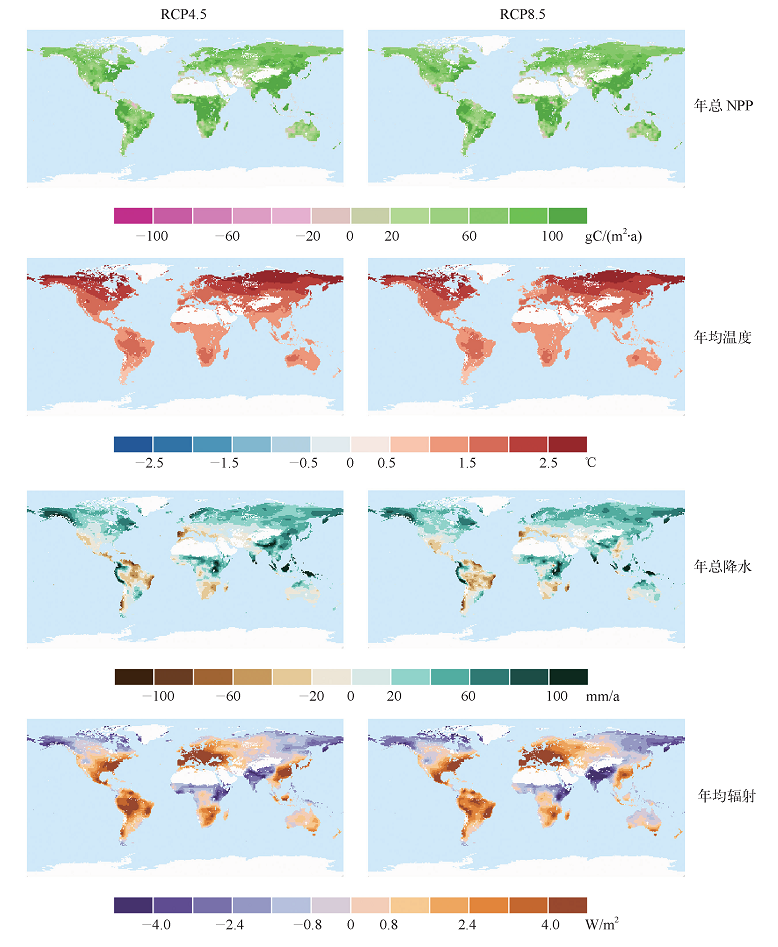 | 图3 全球升温2℃时CMIP5模式在RCP2.6、RCP4.5和RCP8.5情景下预估的全球年总NPP、年均温度、年总降水、年均辐射相对于参考时段(1986— 2005年)的变化量的空间分布Fig. 3 Spatial pattern of changes in annual total NPP, annual mean temperature, annual total precipitation and annual mean down welling shortwave radiation under global warming of 2℃ relative to the reference period 1986 to 2005 based on CMIP5 simulations under RCP2.6, RCP4.5 and RCP8.5 |
2.3 1.5℃和2℃升温情景下各环境因子对NPP变化贡献量的空间格局
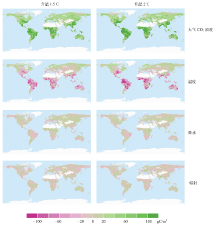
全球尺度的分析表明, 大气CO2浓度增加对陆地生态系统NPP的增加起主导作用, 而温度、降水和辐射对全球NPP的增加贡献量相对较低。本文基于像元尺度进一步分析各环境因子对NPP变化量的贡献量在全球的空间分布。结果表明, 大气CO2浓度和温度的贡献量具有明显的空间格局(图4), 而降水和辐射对NPP变化量的贡献量相对较小。大气CO2浓度和温度对NPP变化的贡献量的空间格局在1.5℃和2℃升温情景下均较为一致, 但升温2℃时各因子的贡献量的绝对值大于升温1.5℃时各因子贡献量的绝对值。升温幅度相同时, RCP2.6、RCP4.5和RCP8.5情景下的各因子的贡献量没有明显区别(未展示)。总体而言, 大气CO2浓度上升对陆地生态系统NPP有促进作用, 大气CO2浓度上升对NPP变化量贡献最大的地区主要分布于中低纬度地区(> 80 gC/(m2· a)), 在高纬度地区贡献量相对较小(20~40 gC/(m2· a))。温度上升对北方高纬度地区和青藏高原地区的NPP增加有促进作用(0~40 gC/(m2· a)), 但对中低纬度地区的NPP具有强烈的抑制作用(< -40 gC/(m2· a))。降水和辐射对NPP变化量的贡献在绝大多数地区均介于-20~20 gC/(m2· a)之间。
 | 图4 全球升温1.5℃和2℃时CMIP5在RCP2.6情景下预估的大气 CO2浓度、温度、降水和辐射对全球年总NPP变化量贡献的空间分布Fig. 4 Spatial pattern of contribution of changes in atmospheric CO2 concentration, temperature, precipitation and down welling shortwave radiation to the changes in annual total NPP under global warming of 1.5℃ and 2℃ based on CMIP5 simulations under RCP2.6 |
针对《巴黎协定》提出的温控目标, 本文利用CMIP5模式在RCP2.6、RCP4.5和RCP8.5情景下的模拟结果初步分析了全球升温情景下陆地生态系统NPP相对于1986— 2005年的变化, 重点讨论了1.5℃和2℃升温情景下NPP的变化量, 并探讨了大气CO2浓度、温度、降水和辐射的变化及其与NPP变化之间的关系。
在未来升温情景下, CMIP5模式在各RCP情景下的模拟结果均表明全球陆地生态系统NPP呈增加趋势, 且NPP增加量与升温幅度成正比。大气CO2浓度、温度、降水和辐射均随着增温幅度的增加而增加。从当前到升温达到《巴黎协定》设定的目标升温幅度时, 全球陆地生态系统NPP总量增加的主因是大气CO2浓度上升, 而温度、降水和辐射的贡献相对较弱。然而, 升温对NPP变化的贡献量具有明显的空间分异:在北方高纬度地区和青藏高原地区, 升温对NPP有促进作用; 在中低纬度地区, 升温对NPP有强烈的抑制作用。北方高纬度地区和青藏高原地区的生态系统NPP主要受温度限制[2], 而这些区域升温速率相对较快[3], 温度的升高从一定程度上缓解了植被的温度限制, 从而促进这些区域NPP的增长。中低纬度地区的升温则可能导致该地区环境温度超过植被生长最适宜温度、降低植被对干旱的抵抗能力, 从而限制生态系统NPP的增长。与升温不同, 大气CO2浓度上升对全球范围内NPP均有促进作用, 且促进作用最大的地区主要分布于中低纬度地区, 可能是由于这些区域内良好的水热条件有利于CO2施肥效应[20, 21]。
本文综合了15个地球系统模式的模拟结果对未来全球升温情景下NPP的相对于1986— 2005年的变化情况进行了分析, 从一定程度上提高了研究结果的鲁棒性。但是值得注意的是, 各个模式的模拟结果之间仍然存在较大的差异。以RCP2.6情景为例, 全球升温1.5℃时, 15个地球系统模式模拟的年总NPP相对于参考时段的增加量为4.17± 2.56 PgC/a, 各模式模拟的年总NPP增加量范围从-0.23 PgC/a (NorESM1-M)至7.92 PgC/a (MPI-ESM-MR)。产生各个模式的模拟结果差异的原因主要包括模式驱动数据集的差异和大气、海洋、陆地模块结构及其耦合方式的差异[3, 22]。相比CMIP4, CMIP5模式考虑了火山喷发和气溶胶的影响, 温度预测性能提高了, 但现阶段地球系统模式仍存在较大的不确定性。例如, CMIP5模式对地球系统内部变率导致的温度变化模拟不足, 且对降水变化趋势的模拟可能存在一致性的高估[22], 这些环境因子模拟的不确定性可能直接导致陆地生态系统NPP模拟的不确定性。此外, CMIP5模式的陆地模块整体仍存在较大的不确定性。就目前的陆地生态系统模型而言, 其模拟NPP的关键生物物理过程中可能存在对CO2施肥效应的高估[23, 24, 25]、对水分条件过于敏感、对营养元素限制、农业管理、土地利用、火灾、臭氧、病虫害等重要过程模拟缺失或不确定性较大[26, 27, 28, 29, 30, 31, 32]。这些问题都需要在未来的研究中不断研究和完善。
本文利用CMIP5模式在RCP2.6、RCP4.5和RCP8.5情景下的模拟结果初步分析了未来全球升温情景下陆地生态系统NPP相对于1986— 2005年的变化量及其可能的驱动因子。然而, 《巴黎协定》的签订意味着本研究所采用的典型浓度路径可能不能代表未来真实情景, 进而可能导致本文模拟的陆地生态系统NPP及其相关环境因子的变化与未来真实情况存在偏差。待《巴黎协定》框架下构建的新的全球温室气体排放路径及其相应的模型模拟结果可获取之后, 有必要对本文的前瞻性的结果进行对比分析。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|